
 Quando si parla di “plasticità” si è soliti riferirsi alla particolare proprietà che ha una sostanza di subire deformazioni di ampiezza significativa e che vengono mantenute nel tempo: come è facilmente intuibile, l’impiego scientifico di questo termine ha trovato ovvia collocazione in discipline come la fisica e l’ingegneria, al fine di descrivere le particolari proprietà meccaniche esibite da alcuni materiali. È stato solo in tempi più recenti che il termine “plasticità” ha trovato una applicazione naturale anche in campo biologico, con riferimento alla modificabilità esibita dai tessuti organici quale risultato dell’esposizione a determinate “esperienze”.
Quando si parla di “plasticità” si è soliti riferirsi alla particolare proprietà che ha una sostanza di subire deformazioni di ampiezza significativa e che vengono mantenute nel tempo: come è facilmente intuibile, l’impiego scientifico di questo termine ha trovato ovvia collocazione in discipline come la fisica e l’ingegneria, al fine di descrivere le particolari proprietà meccaniche esibite da alcuni materiali. È stato solo in tempi più recenti che il termine “plasticità” ha trovato una applicazione naturale anche in campo biologico, con riferimento alla modificabilità esibita dai tessuti organici quale risultato dell’esposizione a determinate “esperienze”.
I fenomeni plastici all’interno del sistema nervoso centrale (SNC) costituiscono il presupposto fondamentale per la formulazione di qualsiasi intervento riabilitativo: essi infatti rappresentano la necessaria premessa teorica, biologicamente fondata, per potere ipotizzare interventi finalizzati a contenere o a superare gli esiti di patologie anche gravi come l’ictus o le lesioni del midollo spinale. Può dunque essere interessante passare in rassegna alcune delle tappe più significative che hanno condotto la comunità scientifica ad accettare l’esistenza della “plasticità neurale” mettendola in relazione con i cambiamenti, talvolta anche notevoli, ai quali va incontro il nostro cervello nel corso della vita per effetto delle esperienze alle quali siamo esposti.
Con le sue osservazioni, Vincenzo Malacarne (1744 – 1816) è stato probabilmente uno tra i primi studiosi a mettere in relazione il fenomeno dell’apprendimento con il tessuto cerebrale. Verso il 1780 egli conduce alcune sperimentazioni impiegando coppie di giovani animali “gemelli” come cani appartenenti alla medesima cucciolata, o pappagalli, cardellini e merli provenienti dalla stessa covata. Il suo scopo dichiarato è quello di stabilire se “l’esercizio continuo e sommamente energico delle facoltà intellettuali non influisca sullo sviluppo primaticcio di alcune parti dell’encefalo”: per questo motivo sottopone uno solo degli animali ad “educazione” ed accerta che le circonvoluzioni cerebrali di quello che era stato avviato al particolare addestramento si sviluppano maggiormente rispetto all’altro.
A conclusioni simili giunse anche il naturalista britannico Charles Darwin (1809 – 1882), noto per la sua teoria dell’Evoluzione delle specie animali per selezione naturale. Nella 1874 Darwin osserva che il volume del cervello del coniglio domestico è inferiore rispetto a quello della lepre e del coniglio selvatico, attribuendo questo fenomeno al fatto che gli animali tenuti in cattività per numerose generazioni non abbiano potuto esercitare l’intelletto, i sensi ed i movimenti volontari.
Le pionieristiche osservazioni di Malacarne e di Darwin rappresentano interessanti anticipazioni del pensiero neuroscientifico successivo: oggi infatti viene ampiamente riconosciuto che la corteccia cerebrale è una struttura estremamente dinamica, e che la sua continua riorganizzazione è da mettere in relazione con le peculiari esperienze compiute. Questa importante acquisizione ha successivamente suggerito la possibilità di individuare specifici metodi di riorganizzazione utili per contenere le conseguenze di lesioni che colpiscono il sistema nervoso centrale (SNC).
Tra la fine del 19° e l’inizio del 20° secolo si assiste ad un rinnovato interesse dei neuroscienziati per i fenomeni di degenerazione e rigenerazione cellulare che avvengono entro il SNC. Il termine “plasticità neuronale” comincia così a far parte del vocabolario impiegato dai ricercatori nelle comunicazioni scientifiche e nel 1896 compare nel titolo della pubblicazione del belga Jean Demoor (1867 – 1941) “La plasticité morphologique des neurones cérébraux”.
Tuttavia, restava ancora molto da scoprire sui meccanismi effettivi che soggiacciono alla plasticità, anche perché a quell’epoca la comunità scientifica internazionale dibatteva ancora aspramente su quale fosse la “struttura” del tessuto cerebrale e, conseguentemente, rimaneva oscura la modalità di trasmissione dell’impulso nervoso: i due principali filoni di pensiero contrapposti erano rappresentati dall’italiano Camillo Golgi (1843 – 1926) e dallo spagnolo Santiago Ramon y Cajal (1853 – 1934). Golgi era il principale sostenitore della “teoria reticolare”, che concepiva il sistema nervoso come un insieme continuo ed ininterrotto di tutti i suoi elementi. Al contrario, Cajal era il principale esponente della “teoria del neurone”, che prevedeva che i neuroni, separati gli uni dagli altri, potessero comunicare con quelli contigui attraverso zone specializzate chiamate “sinapsi”. Per una descrizione dei principali elementi che costituiscono un neurone potete guardare la Fig. 1 qui di seguito.

Fig. 1. Disegno schematico di un neurone
È in questo contesto che sin dal 1893 l’italiano Eugenio Tanzi (1856 – 1934), aderendo alla “teoria del neurone” proposta da Cajal, intuì che lo stimolo nervoso doveva incontrare una qualche resistenza nel passare da un neurone a quello successivo. Per spiegare il fenomeno dell’apprendimento, egli suppose che per effetto della stimolazione ripetuta si riducesse la distanza tra i neuroni funzionalmente connessi, dunque facilitando il passaggio dell’eccitazione nervosa: ciò sarebbe stato possibile solo se i neuroni fossero stati in grado di andare incontro a cambiamenti morfologici e dunque Tanzi ipotizzò che le arborizzazioni terminali dell’assone di un neurone che si trovava “a monte” dovessero potere crescere in lunghezza, avvicinandosi ai dendriti di quello situato “a valle” (Berlucchi, Butchel, 2009).
Influenzato da Tanzi, del quale era stato allievo, anche il medico e psichiatra italiano Ernesto Lugaro (1870 – 1940) ha dato contributi fondamentali allo studio della plasticità neurale. Nel 1906 egli pubblicò un trattato di Psichiatria in cui fenomeni come l’apprendimento e la compensazione dei deficit funzionali venivano attribuiti alla plasticità neuronale. Da alcuni, Lugaro viene considerato come il primo dei neuroscienziati ad impiegare esplicitamente il termine “plasticità” in una accezione molto simile a quella moderna, con riferimento non soltanto alla modificabilità morfologica del neurone ma anche intuendo l’importanza ricoperta dai processi chimici ai fini della trasmissione sinaptica (Berlucchi, 2002).
Più o meno negli stessi anni, nel 1909, il medico rumeno Ioan Minea (1878 – 1941) pubblica una tesi dal titolo “Investigazioni sperimentali sulle variazioni morfologiche dei neuroni sensoriali (studi sulle “reazioni plastiche”)”. È molto probabile che Minea avesse ripreso il termine “plasticità neuronale” dal suo maestro Georges Marinesco (1863 – 1938), che viene considerato come il fondatore della neurologia clinica romena moderna. Minea amplia gli studi di Marinesco sul trapianto delle cellule gangliari e descrive una serie di modificazioni morfologiche alle quali va incontro questa cellula nervosa: tra queste, la “rigenerazione collaterale”, la “plasticità dendritica” (estensione di nuovi processi) e la “plasticità (proliferazione) di neurofibrille (Jones, 2000).
Anche nel titolo di due distinti lavori pubblicati nel 1925 e nel 1931 dal fisiologo tedesco Albrecht Bethe (1872 – 1955) si trova il termine “plastizität“: l’autore si riferisce ad essa come alla capacità esibita da un organismo di adattarsi ai cambiamenti interni ed esterni attraverso la mediazione del SNC. Tale capacità di adattamento del SNC veniva presa in considerazione anche quale mezzo per fare fronte a lesioni più o meno estese, anche se veniva concepita più nei termini di una riorganizzazione del tessuto rimasto integro e non tanto come una risposta che coinvolgeva il livello neuronale (Pia, 1985).
Nonostante le numerose anticipazioni, in molti ritengono che il neurofisiologo polacco Jerzy Konorski (1903 – 1973) debba essere considerato come lo studioso grazie al quale la “plasticità neuronale” ha fatto autorevolmente il suo ingresso nelle Neuroscienze. Nella sua importante monografia “Conditioned reflexes and neuron organization” pubblicata nel 1948 egli non soltanto definisce il concetto di plasticità neuronale, ma avanza ipotesi sui meccanismi che la sottendono, e che secondo lui sono da ricercare nella riorganizzazione delle connessioni sinaptiche (Konorski, 1948). Konorski considera la plasticità neuronale come una proprietà fondamentale del SNC degli animali superiori, che può costituire il meccanismo comune per processi quali l’apprendimento, la memoria, lo sviluppo cerebrale ed il recupero che si osserva dopo una lesione al cervello (Butchel, 1978).
La questione riguardante la possibilità del tessuto nervoso di riorganizzarsi dal punto di vista anatomico continuò ad appassionare il dibattito neuroscientifico, e molti altri ricercatori cominciarono sempre più a ritenere che le modificazioni plastiche neuronali potessero costituire il substrato materiale di processi quali la memoria, l’apprendimento e il recupero che si osserva dopo una lesione. Tuttavia, mentre i processi di rigenerazione nervosa periferica erano relativamente più semplici da spiegare, quelli che avevano luogo entro il sistema nervoso centrale continuavano a presentare numerose difficiltà interpretative. Infatti, se viene leso un nervo periferico, come nel caso dell’interruzione dell’assone di un neurone di moto proveniente dal midollo e che raggiunge alcune fibre muscolari, la rigenerazione delle fibre interrotte può, virtualmente, condurre al ripristino delle connessioni originali (vedi Fig. 2).
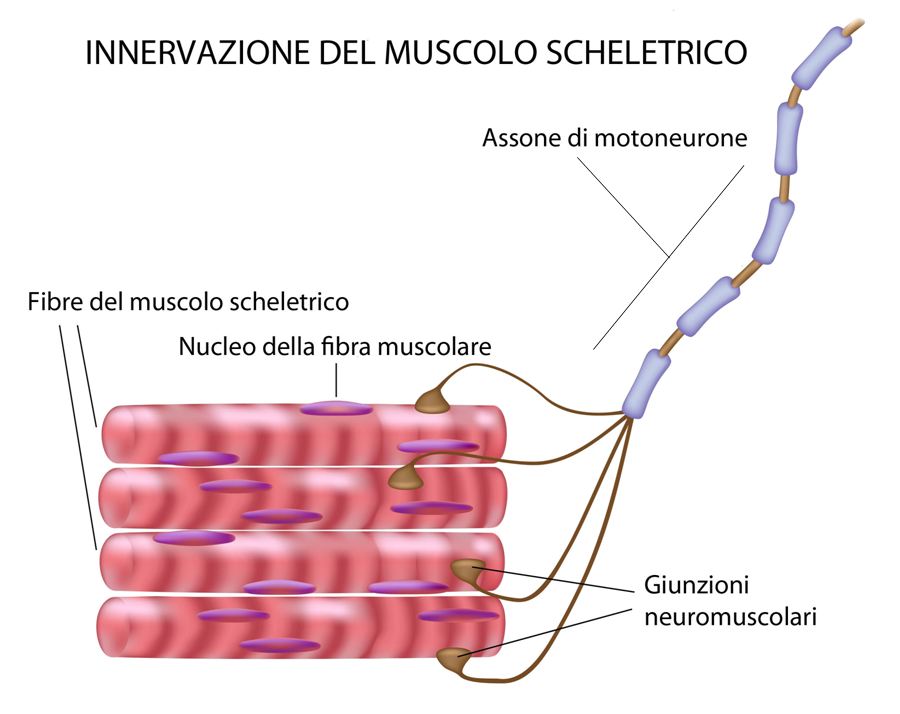
Fig. 2. Nel sistema nervoso periferico, la rigenerazione di un assone che è stato leso può determinare la ricostituzione dei contatti con le giunzioni neuromuscolari originari
Entro il sistema nervoso centrale, la situazione è molto diversa poiché questa modalità di rigenerazione non è parsa possibile sin dalle prime ricerche dell’epoca sull’argomento. Tuttavia i ricercatori hanno cominciato ad ammettere l’esistenza di una seconda possibilità di rimodellamento post-lesionale, che consiste nella crescita di nuovi terminali che partono dalle zone adiacenti alla lesione e vanno a reinnervare le aree rimaste deafferentate entro il cervello o entro il midollo spinale. Si tratta delllo sprouting collaterale (vedi oltre), fenomeno che ha posto sin dall’inizio la questione sulla “specificità” delle connessioni di nuova formazione: si sospettava infatti che una reinnervazione eterogenea e scarsamente selettiva entro un ambiente come il sistema nervoso centrale, più “caotico” rispetto al sistema nervoso periferico (vedi Fig. 3) avrebbe potuto avere uno scarso significato funzionale anche dal punto di vista della qualità del recupero (Raisman, 1969).
Fig. 3. Reti neurali all'interno del Sistema Nervoso Centrale
Anche se l’interpretazione dei fenomeni plastici post-lesionali lasciava spazio a dubbi che, come vedremo, perdurano anche ai nostri giorni, le ricerche che interessavano soggetti integri producevano ulteriori conoscenze. Di grande interesse sono alcuni studi che hanno messo in relazione le modificazioni a cui va incontro il SNC di alcuni animali da laboratorio a seconda del differente ambiente in cui sono stati costretti a vivere. Agli inizi degli anni ’70 del secolo scorso, riprendendo le osservazioni che Vincenzo Malacarne aveva compiuto due secoli prima, il neuroscienziato Mark Richard Rosenzweig (1922 – 2009) rilevava che vi erano notevoli differenze tra i cervelli dei roditori che venivano allevati in un ambiente “arricchito” e quelli dei loro simili che venivano mantenuti in gabbie “standard”. I ratti che vivevano insieme ad altri in gabbie ricche di stimoli e che, per nutrirsi, dovevano percorrere labirinti o risolvere svariati “problemi” sviluppavano maggiormente la corteccia cerebrale rispetto ad altri animali che, al contrario, erano stati mantenuti in condizioni di isolamento ed in un ambiente monotono.
Queste ricerche sono molto significative non soltanto perché confermano le conseguenze più macroscopiche prodotte dall’esperienza sul tessuto cerebrale come l’aumento del volume e del peso del cervello, ma anche perché forniscono una serie di interessanti dati microanatomici. Infatti, viene qui dimostrato che i neuroni cerebrali dei ratti esposti all’ambiente arricchito risultano maggiormente interconnessi grazie ad un aumento della densità della loro ramificazione dendritica, e che l’addestramento altera i processi elettrochimici migliorando l’efficacia della trasmissione sinaptica (Rosenzweig, Bennett, Diamond, 1972).
Particolarmente interessanti sono anche gli studi di Patrick David Wall (1925 – 2001) . Egli osserva che in genere il recupero è un fenomeno particolarmente lento, e che, allo stesso modo, il fattore tempo è di importanza cruciale anche per la manifestazione dell’insorgenza di un danno a carico del SNC. Ad esempio, è noto che le lesioni che distruggono lentamente il tessuto cerebrale, come i tumori cerebrali, solitamente danno luogo a sintomi evidenti solo quando raggiungono una dimensione rilevante: a parità di estensione, tuttavia, lesioni improvvise come l’ictus, producono istantaneamente danni gravi e manifesti. Secondo Wall, questa constatazione è di per sé sufficiente ad ammettere l’esistenza di una qualche forma di processo di recupero all’interno del SNC, che potrebbe compensare, entro certi limiti, l’effetto prodotto dalle lesioni che insorgono progressivamente ed in tempi lunghi.
Wall inoltre invoca due principali e distinti meccanismi implicati nel recupero: lo “smascheramento di sinapsi” e i “meccanismi plastici” propriamente detti. Egli innanzitutto premette che le cellule nervose mostrano un comportamento definibile “omeostatico” per effetto del quale, nel periodo successivo ad una lesione, è presente una plasticità riorganizzativa massiva. Quando al tessuto nervoso rimasto integero viene a mancare una parte di connessioni, come accade nel caso in cui parte dell’input viene meno perché proveniva da altri neuroni che sono stati lesionati, si assiste ad un iniziale periodo di inibizione (vedi anche “diaschisi“) al quale fa seguito una fase di massima plasticità in cui i neuroni superstiti che sono rimasti deprivati del loro normale flusso afferente tendono ad aumentare la propria eccitabilità in maniera tale da ricostituire la loro quota di contatti sinaptici.
Nella Fig. 4 viene mostrato schematicamente questo concetto. Nella parte sinistra della figura si vede che il neurone “A” è collegato a “B” mentre “C” è collegato a “D” per mezzo di connessioni che sono rappresentate dalle frecce: la linea “x“, più sottile, rappresenta una connessione tra “A” e “D” che tuttavia rimane “nascosta”, poiché l’organismo non le attribuisce un significato funzionale rilevante. Wall sottolinea infatti che con il processo di maturazione dell’individuo, molte delle connessioni inutili tra i neuroni presenti alla nascita vengono eliminate o represse. A destra della figura, il neurone “C” è andato distrutto dalla lesione, così come il suo prolungamento assonale verso “D“: con il simbolo “!” viene indicato il sito sinaptico lasciato vuoto dalla fibra degenerata.

Fig. 4. Se una lesione colpisce il neurone "C" (a sinistra del disegno), questi andrà incontro a degenerazione, e con lui il prolungamento assonico che precedentemente occupava il sito sinaptico sul neurone "D" (a destra del disegno)
Nella Fig. 5, a sinistra viene mostrato il meccanismo riorganizzativo post-lesionale sostenuto dallo “smascheramento sinaptico“, per effetto del quale la connessione “x“, precedentemente silente, entra ora in azione. A destra viene invece mostrato il caso in cui il rimodellamento plastico ha luogo, in un tempo maggiore, per effetto del fenomeno dello “sprouting“, ovvero della gemmazione di nuove arborizzazioni assoniche ad opera di neuroni rimasti risparmiati dalla lesione, indicata dalla freccia tratteggiata contrassegnata dalla lettera “y“.

Fig. 5. A sinistra viene mostrato il meccanismo dello "smascheramento sinaptico" per il quale, connessioni precedentemente silenti diventano attive. Questo processo viene invocato per spiegare le modificazioni plastiche molto veloci che insorgono a breve distanza di tempo dalla lesione. A destra viene mostrato il meccanosmo dello "sprouting collaterale", per effetto del quale, arborizzazioni di nuova formazione vanno ad occupare il sito sinaptico vacante del neurone "D".
Wall distingue dunque lo “smascheramento di sinapsi” dalle modificazioni plastiche vere e proprie, consistenti nell’emissione di assoni completamente nuovi. In particolare, lo smascheramento di sinapsi può spiegare quei rimodellamenti neurali che si verificano in tempi troppo rapidi per potere essere spiegati con la plasticità vera e propria o “sprouting”, che richiederebbe tempi molto più lunghi. Nella sua lucida esposizione, tuttavia, Wall non manca di rilevare che lo smascheramento di sinapsi non rappresenta necessariamente un meccanismo favorevole al migliore recupero possibile: proprio perché si tratta di collegamenti che erano stati “inattivati” perché ritenuti poco rlevanti, essi potrebbero divenire veicolo di informazioni prive di senso dal punto di vista funzionale (Wall, 1983).
Occorre qui sottolineare che anche alla plasticità neurale in quanto tale non è sempre stato attribuito un significato intrinsecamente positivo. Questo sospetto ha trovato conferme in alcune ricerche che risalgono al 1958, che hanno fornito le prove elettrofisiologiche ed istologiche del fatto che gli assoni delle radici dorsali del midollo possono generare nuovi terminali andando ad occupare i siti terminali che sono stati lasciati vuoti sia per degenerazione degli assoni della radice dorsale adiacente come pure per degenerazione dovuta a sezione dei tratti discendenti, cosa che effettivamente ha luogo in caso di lesione midollare o cerebrale. Dal punto di vista clinico, dunque, quando i terminali delle fibre discendenti degenerano in seguito a lesione, essi possono essere rimpiazzati da nuovi sprouting provenienti dalle fibre afferenti midollari vicine: in questo modo esse acquisiscono una anomala prevalenza sugli archi riflessi, e divengono causa della “spasticità” (vedi anche “lo specifico motorio nel paziente emiplegico“) o, almeno, dell’aumento del riflesso allo stiramento (McCouch, Austin, Liu, Liu, 1958).
Quest’ultimo aspetto presenta importanti implicazioni anche dal punto di vista riabilitativo. Wall aveva infatti descritto la spiccata tendenza del tessuto neurale risparmiato da una lesione a ricostituire i contatti sinaptici perduti: indubbiamente questa proprietà neuronale crea una condizione per la quale il periodo post-lesionale è caratterizzato da una estrema plasticità che, tuttavia, in alcuni casi potrebbe anche essere maladattiva. Era già stato infatti sperimentalmente dimostrato che ha luogo una sorta di ”competizione sinaptica” tra i neuroni integri per andare ad occupare i terminali sinaptici rimasti vacanti, fenomeno che potrebbe anche condurre alla formazione di connessioni nuove ma anomale: in questa “competizione” sarebbero favoriti i neuroni topograficamente più vicini e non necessariamente quelli più “adatti” dal punto di vista funzionale (Fig. 6) (Schneider, 1973).

Fig. 6. Nel periodo post-lesionale si assiste ad una "competizione" per l'occupazione dei siti sinaptici vacanti. È stato fatto osservare come, tale processo posa anche essere alla base di fenomeni plastici maladattivi
Questo concetto è stato ripreso anche più recentemente, con l’osservazione che, ad esempio, lesioni cerebrali che avvengono nel primo periodo di vita possono stimolare importanti riorganizzazioni anatomiche che potrebbero rivelarsi controproducenti per l’acquisizione delle abilità necessarie in un periodo successivo (Gardner, 1983).
Questi studi hanno una notevole importanza anche dal punto di vista riabilitativo, poiché permettono di concepire il sistema nervoso come un tessuto estremamente plastico, suscettibile di rimodellamento anche notevole sia per l’esposizione a determinate esperienze, sia in seguito a lesione; di conseguenza, cominciava a farsi strada la necessità di un intervento di tipo “tecnico” e precoce su questi processi “naturali” al fine di ottenere una riorganizzazione dei sistemi lesi con caratteristiche qualitative migliori di quelle che avrebbero luogo spontaneamente. Nel soggetto sano l’acquisizione di nuove competenze motorie, cognitive, linguistiche, ecc., si riflette nelle modificazioni a carico del tessuto nervoso che fa seguito all’esposizione a determinate “esperienze”: il “recupero” è il nome con il quale la Riabilitazione Neurocognitiva designa processi, in parte simili, di “apprendimento” che hanno luogo in condizioni patologiche (Perfetti, 1979).
Tra la fine degli anni ’70 e gli inizi degli anni ’80 del secolo scorso vennero pubblicati numerosi lavori che fornivano chiare indicazioni del fatto che le aree (o “mappe”) nelle quali siamo soliti suddividere rigidamente la corteccia cerebrale sono in realtà molto dinamiche, variano in ampiezza e modificano i propri confini continuamente a seconda delle esperienze che facciamo e, cosa importante, ciò avviene anche nella vita adulta e non solo nell’organismo in via di sviluppo. A testimonianza del grande interesse per l’argomento, nel 1982 il termine “neuronal plasticity” entra a fare parte delle “MESH”, che è una sorta di vocabolario dei termini sulla base dei quali Pubmed, che è un autorevole motore di ricerca degli articoli scientifici di tutto il mondo, cataloga le pubblicazioni. Il concetto di “plasticità” era ormai entrato a pieno titolo nel dibattito scientifico e cominciarono ad uscire sempre più numerose pubblicazioni riguardanti le ampie possibilità di riorganizzazione post-lesionale nel soggetto adulto e non solo, come si tendeva ancora talvolta a ritenere, in quello in età evolutiva.
In questo senso, possono essere interessanti le ricerche sulle modificazioni che si vengono a creare nella corteccia sensoriale primaria della scimmia adulta in seguito alla interruzione di un nervo periferico che innerva la mano. Se viene leso il nervo mediano, le informazioni sensoriali cutanee provenienti da alcune dita non possono raggiungere l’area di rappresentazione corticale corrispondente: tuttavia, dopo qualche tempo, l’area di rappresentazione corticale del nervo mediano non rimarrà silente, ma verrà occupata dalle rappresentazioni delle altre dita della mano che sono innervate dal nervo radiale e dal nervo ulnare rimasti integri. Cosa interessante, i campi recettivi delle dita rimaste innervate divengono più piccoli (più il campo recettivo è piccolo e maggiore è la sensibilità cutanea) quando le loro aree di rappresentazione corticale si sono ampliate a spese dell’area di rappresentazione del nervo mediano. Queste ricerche mostrano dunque che anche nell’adulto le aree di rappresentazione corticale non esprimono una cartografia statica: al contrario, la loro organizzazione è estremamente dinamica e riflette l’esperienza del soggetto o, come in questo caso, la sua deprivazione (Merzenich et al., 1983).
Oltre alle ricerche che hanno messo in evidenza le riorganizzazioni, talvolta massive, che fanno seguito ad una lesione, oggi possiamo disporre di studi che dimostrano la stretta correlazione tra modificazioni plastiche del tessuto nervoso ed acquisizione di una determinata abilità motoria, con evidenti suggestioni anche per la Riabilitazione. Ad esempio, scimmie adulte addestrate a riconoscere uno stimolo tattile esibiscono un progressivo miglioramento della loro abilità, che è stato spiegato con due processi differenti ma interconnessi: i progressi iniziali sono stati attribuiti a fattori “cognitivi” quali l’attenzione al compito e l’adozione di nuove strategie, mentre quelli a comparsa più tardiva sono stati interpretati come il prodotto di “modificazioni strutturali neurali plastiche” delle aree di rappresentazione delle dita coinvolte nell’azione richiesta (Recanzone et al. 1992).
E proprio la stretta relazione tra i processi cognitivi e la qualità delle riorganizzazioni plastiche del tessuto nervoso che si accompagna alla acquisizione di nuove abilità nell’organismo sano suggeriscono quali possano essere le strategie di intervento riabilitativo finalizzate a minimizzare o a contenere gli effetti di una lesione: “recupero” è infati il nome che può essere dato a quei processi di “apprendimento” che hanno luogo in condizioni patologiche, secondo quanto proposto dalla Teoria Neurocognitiva della Riabilitazione.
La dimostrazione del fatto che i processi cognitivi siano fondamentali nell’acquisizione di nuove abilità proviene da numerose ricerche, che è impossibile anche solamente elencare: una, ad esempio, ha confrontato l’espansione della corteccia motoria primaria di scimmie adulte che eseguivano compiti solo apparentemente simili. Scimmie che dovevano estrarre del cibo da piccoli buchi hanno mostrato un incremento della loro abilità motoria che si è accompagnata ad una espansione della rappresentazione dei movimenti della parte distale del’arto impiegato nel compito. Viceversa, un secondo gruppo di scimmie, al pari delle altre, aveva ripetutamente estratto del cibo da buchi più grandi, senza avere avuto la necessità di sviluppare alcuna particolare strategia motoria: in questo caso, non si è verificata alcuna modificazione dell’area di rappresentazione dei movimenti dell’arto (Plautz, et al., 2000).
Questi recenti dati sperimentali mettono in evidenza il fatto che, di per sé, la ripetizione del movimento non genera alcuna modificazione funzionale delle mappe corticali, confermando l’ipotesi che, lo sviluppo di una abilità consiste, piuttosto, nella ripetizione del processo di soluzione di un determinato problema motorio (Bernštein, 1966).
In conclusione, si possono fare alcune considerazioni in chiave riabilitativa. Innanzitutto, esistono numerose evidenze del fatto che il tessuto nervoso va incontro a riorganizzazioni anche notevoli in seguito ad una lesione: tuttavia, questi rimodellamenti plastici non sempre costituiscono il substrato per il migliore recupero della funzione lesa, e talvolta sono alla base di fenomeni patologici come la spasticità, già descritta in precedenza, o l’insorgenza dell’arto fantasma (Ramachandran, Hirstein, 1998).
Per questo motivo, alla plasticità neurale non è possibile attribuire un significato finalistico intrinsecamente positivo: essa piuttosto rappresenta una opportunità, che può essere sfruttata per modificare l’evoluzione per così dire “naturale” di un evento lesivo. La modificabilità biologica del tessuto nervoso costituisce pertanto il presupposto fondamentale per interventi guidati dal Terapista, che persegue lo scopo di favorire riorganizzazioni migliori di quelle che avrebbero luogo spontaneamente. Nella Teoria Neurocognitiva della Riabilitazione l’esercizio viene ad essere concepito come la risoluzione di un particolare “problema” (vedi anche questo link) che il malato deve risolvere attivando processi cognitivi quali l’Attenzione, la Percezione, la Memoria, ecc., in un’ottica in cui il recupero è un processo di Apprendimento che avviene in condizioni patologiche (vedi anche questo link).
BIBLIOGRAFIA
Berlucchi G, Butchel HA. Neuronal plasticity: historical roots and evolution of meaning. Exp Brain Res, 2009: 192(3): 307-19;
Berlucchi G. The origin of the term plasticity in the Neurosciences: Ernesto Lugaro and chemical synaptic transmission. J Hist Neurosci, 2002: 11(3): 305-309;
Bernštein NA. La coordinazione dei movimenti nell’ontogenesi (ed. orig. 1966). In: Bernštein NA, “Fisiologia del movimento”, Roma: S.S.S. Ed.; 1989;
Butchel HA. On defining neural plasticity. Arch Ital Biol, 1978: 116(3-4):241-7;
Gardner H. Formae mentis. Saggio sulla pluralità dell’intelligenza. Milano: Feltrinelli; 1987
Konorski J. Conditioned reflexes and neuron organization. Cambridge University Press, 1948;
Jones EG. Plasticity and Neuroplasticity. J Hist Neurosci, 2000: 9(1): 37-39;
McCouch GP, Austin GM, Liu CN, Liu CY. Sprouting as a cause of spasticity. J Neurophysiol, 1958: 21(3):205-16;
Merzenich MM, Kaas JH, Wall J, Nelson RJ, Sur M, Felleman D. Topographic reorganization of somatosensory cortical areas 3b and 1 in adult monkeys following restricted deafferentation. Neurosci 1983: 8(1): 33-55;
Perfetti C. La rieducazione motoria dell’emiplegico. Milano: Ghedini Ed.; 1979;
Pia HW. Plasticity of the central nervous system: a neurosurgeon’s experience of cerebral compensation and decompensation. Acta Neurochir, 1985: 77(3-4): 81-102;
Ramachandran VS, Hirstein W. The perception of phantom limb. Brain, 1998: 121(9): 1603-30;
Recanzone G, Jenkins WM, Hradek GT, Merzenich MM. Progressive improvement in discriminative abilities in adult owl monkeys performing a tactile frequency discrimination task. J Neurophysiol 1992: 67(5): 1015-30;
Plautz EJ, MillikenGW, Nudo RJ. Effects of repetitive motor training on movement representations in adult squirrel monkeys: role of use versus learning. Neurobiol Learn Mem, 2000: 74(1):27-55;
Raisman G. Neuronal plasticity in the septal nuclei of the adult rat. Brain Res,1969: 14(1): 25-48;
Rosenzweig MR, Bennett EL, Diamond MC. Brain changes in response to experience. Sci Am 1972: 226(2): 22–29
Schneider GE. Early lesions of superior colliculus: factors affecting the formation of abnormal retinal projections. Brain Behav Evol, 1973: 8(1): 73-109;
Wall PD. Meccanismi della plasticità di connessione in seguito ad un danno nel sistema nervoso centrale. In: (a cura di) Paul Bach-y-Rita, “Recupero funzionale”, Cosenza, Brenner Ed., 1983;
In base all’articolo desidererei sapere se:è prevista la possibilità che a distanza di quattro anni dall’evento che mi ha colpito (ictus emorragico con esiti di emiplegia sx.)la plasticità cerebrale potrà ancora sopperire ai circuiti neuronali andati “persi” e se possa esistere un collegamento con la “Nutraceutica”. Grazie per la risposta: Albertini Luigi, età 50 anni,Verona.
Caro Andrea,
è tanto che non ci si sente e spero tutto bene voi.
Da parte mia tutto ok.
Come vedi l’interesse per la riabilitazione neurocognitiva non si esaurisce mai. Non so se hai saputo che ora nel mio studio collabora anche il Dr Fabio Conti e ne sono felice.
Grazie e complimenti per avermi dato l’opportunità di leggere io tuo interessantissimo articolo.
Un caro saluto
Alberto Ghirlanda
Dr med. Alberto Ghirlanda
Specialista FMH Medicina interna generale
CH-6532 Castione / TI
T +41 91 829 33 33
C +41 91 829 44 44
N +41 79 620 30 50
email – ghirla@hotmail.com